

“Sing like you are holding an egg in your mouth.”
“Stretch your phrases like you are pulling taffy.”
“Imagine you are a puppet, with a string attached to your head.”
If you have never taken a music lesson in your life, these instructions probably seem a little silly to you. For those of you who have, you have probably had a teacher use some fairly abstract images or metaphors like these at one point or another to influence your sound or playing technique. These tricks are especially common amongst voice teachers, as they must teach their students how to manipulate tiny internal muscles and structures, without being able to see them, in order to produce a beautiful sound. Victor Fields (1972:2) claims, “vocal expression is…governed by the singer’s powers of visualization and mental imagery” and, because of this, that “we must learn to sing in thought, for the tone is embedded in the idea that produces it.” By using imagery in a teaching setting, the teacher is able to tailor the lessons to the student’s understanding. This metaphorical approach contrasts strongly with a purely physiology-based teaching method, and has prompted several debates over which is more effective. More recently, it has come to light that the most effective method may, in fact, be the result of combining the two approaches. This idea of combining the scientific and the figurative parts of vocal pedagogy lead me to wonder: what if we could understand the biological basis behind how these metaphors and images are used? Would it lead to a better way to tailor teaching methods to students? Let us first investigate further this idea of combining scientific and abstract teaching methods.
[accordion multiopen=”true”]
[toggle title=”In the brain or in the body: Is a scientific approach better than using abstract images and metaphors?” state=”opened”]
There are, however, many that believe a purely physiological approach is best when it comes to teaching, claiming, like Miller (1998:41-42), that the metaphorical examples are merely “useless admonitions” and “illogical.” The field of vocal science has continued to evolve since its foundation by Manuel Garcia (Clements, 2008), and as it does, growing numbers of voice  teachers are leaving behind metaphorical language for a more literal and technical explanation of how the voice works. Richard Miller (2001:49) is a firm believer in this science-based approach, stating that it is “far easier, far simpler, far faster [and] far more honest. Arguments for this type of thinking include the fact that imagery was not used historically and that figurative language is not vital or understandable.
teachers are leaving behind metaphorical language for a more literal and technical explanation of how the voice works. Richard Miller (2001:49) is a firm believer in this science-based approach, stating that it is “far easier, far simpler, far faster [and] far more honest. Arguments for this type of thinking include the fact that imagery was not used historically and that figurative language is not vital or understandable.
Just as imagery might be hard to understand for some, scientific information can also be hard to process. Both approaches have the potential to be unsuccessful if there is a lack of understanding, either on the part of the teacher or the student. Since it is apparent that different students require different teaching methods, the most logical solution is to combine a variety of different methods. William Vennard (1967:147) supports this, claiming that “scientific language is inadequate in teaching an art, and we fill out the deficiency with poetic imagery.” From this point of view, a teacher would have to be able to understand and convey the basic physiological principles, while using imagery to reinforce and unify those concepts and to allow students to get beyond the notes to the emotional subtext. Just as we need a physiological basis on which to this build metaphorical teaching method, a basic understanding of neural anatomy is the first step in understanding how we process the metaphors.
[/toggle]
[toggle title=”Building blocks of language: what are the areas of the brain involved in language processing?”]
In order to be able to locate the areas of the brain that may be activated during metaphor processing, we must first understand the layout of the brain. The central nervous system, which consists of the brain and the spinal cord, are made up of seven basic parts: the spinal cord, the medulla, the pons, the cerebellum, the midbrain, the diencephalon and the cerebrum (Purves et al., 2008).
The most prominent of the structures are the cerebral hemispheres, the left and right sides if the cerebrum. They are characterized by gyri (singular: gyrus), which are the crests of the continuous, highly folded patterns that make up the outer portions of the cortical tissue, and sulci, the grooves that divide the gyri from each other. The patterns of gyri and sulci vary from person to person, but the cerebral hemispheres can still be divided into consistent parts, known as lobes. These are the occipital, temporal, parietal, and frontal lobes, named for the bones of the cranium that protect each of them (Purves et al., 2008).
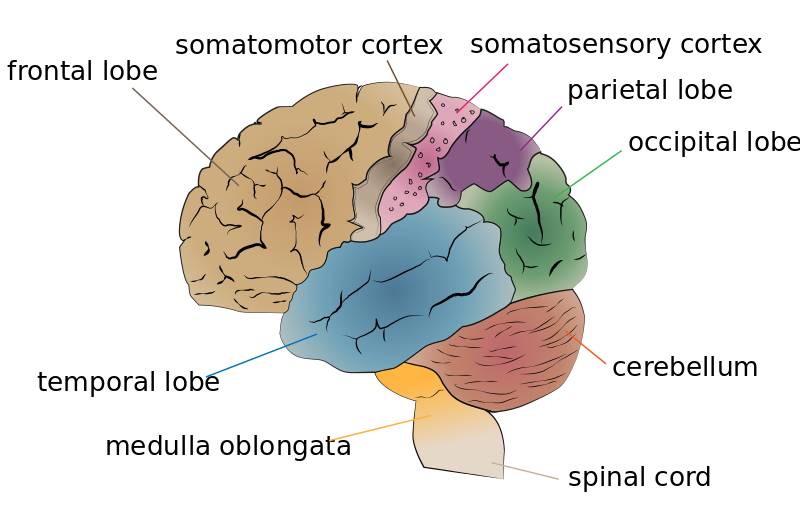
As you can see on the previous diagram, two of the central features of the frontal and parietal lobes are the motor cortex and the sensory cortex. These structures are responsible for housing the neurons that control motor behaviours and process sensory information, respectively (Purves et al., 2008). These areas are just a couple of the specialized areas of the brain, which you can see in more detail here:
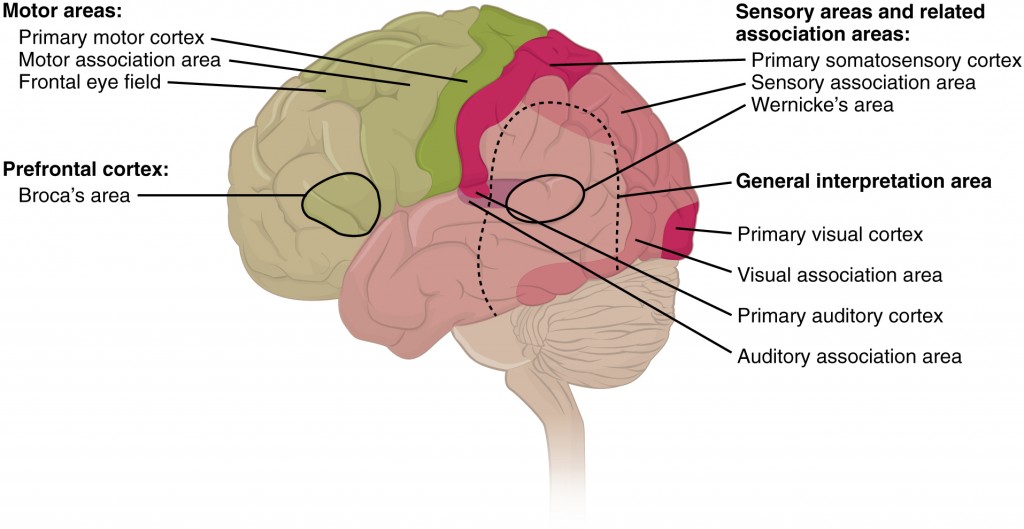
Scientists have located two areas of the brain that are vital for language processing: Broca’s area and Wernicke’s area. Broca’s area is located in the left frontal lobe and is responsible for language production, whereas Wernicke’s area is a region of cortex located in the superior and posterior region of the left temporal lobe. Wernicke’s area helps in mediating language comprehension, creating links between speech sound and meaning that then interact with the circuitry of Broca’s area to enable motor commands that organize the production of meaningful speech (Purves et al., 2008). Armed with a better idea of where the specialized areas of the brain are located and what their individual functions are, we can now begin to look at how these areas might be involved in metaphor processing.
If you find yourself needing more information about the anatomy of the brain, feel free to watch this series of videos, created to explain each part of the brain in detail.
[/toggle]
[toggle title=”Mapping the brain: which areas of the brain are activated while processing auditory and visual cues?”]
In processing metaphors, the brain must first have some sensory input as a starting point, which is usually an auditory or visual cue. In the case of an auditory cue, sound waves are collected by the outer ear and are channeled through the inner structures of the ear, creating a signal that activates the auditory nerve. The information is then carried to the central nervous system, more specifically the auditory cortex, located in the temporal lobe. Visual cues are processed similarly, with photoreceptor cells turning light energy into electrical energy that travels along the optic nerve to the visual cortex, found in the posterior region of the occipital lobe (Lane, 2002).
 According to Stephen Kosslyn (1994: 287), “images are formed by the same processes that allow one to anticipate what one would see if an object were actually present.” Following this logic, a metaphorical image would be created in the brain through the same pathways regardless of whether the incoming cue was auditory or visual, with the exception of the language processing centers. Whether the student was shown a visual representation of what they were being asked to do, or the image was merely described to them, it is thought the image would be created or processed by the same regions of the brain.
According to Stephen Kosslyn (1994: 287), “images are formed by the same processes that allow one to anticipate what one would see if an object were actually present.” Following this logic, a metaphorical image would be created in the brain through the same pathways regardless of whether the incoming cue was auditory or visual, with the exception of the language processing centers. Whether the student was shown a visual representation of what they were being asked to do, or the image was merely described to them, it is thought the image would be created or processed by the same regions of the brain.
[/toggle]
[toggle title=”Left-brain or right-brain: which side of the brain controls the voice during singing?”]
We have all seen those quizzes that tell us which side of our brain is dominant. This dominance refers lateralization of the brain, or the lateral division of the brain into two hemispheres, each of which has functional specializations that are not necessarily performed in the other.
In the majority of people, the primary language functions for spoken language processing are located in the left hemisphere of the frontal and temporal lobes. This is due to a left-sided predominance of the grammatical and syntactical aspects of language. This dominance of the left-side leads to the misguided belief that it is completely responsible for speech production, when, in fact, the right-side governs the emotional context of speech (Purves et al., 2008).
When looking at this lateralization effect in vocal music performance, it has been found that there is a much less pronounced lateralization effect than seen in speech production. However, it appears that during vocal music performance, the right-side is more highly activated. This was true for both physical situations and imagined situations, such as mental practice. (Ackermann, Wildgruber & Riecker, 2006)
[/toggle]
[toggle title=”Continuing the path: how does the brain control vocal production?”]
As mentioned above, there have been studies done to determine which areas of the brain are activated during both overt and covert (real and imagined) singing and speech in the brain. In the case of overt singing and speech, both motor cortices were activated, with the right cerebellum and insula during singing and the left cerebellum and insula during speech (Riecker, Ackermann, Wildgruber, Dogil & Grodd, 2000).
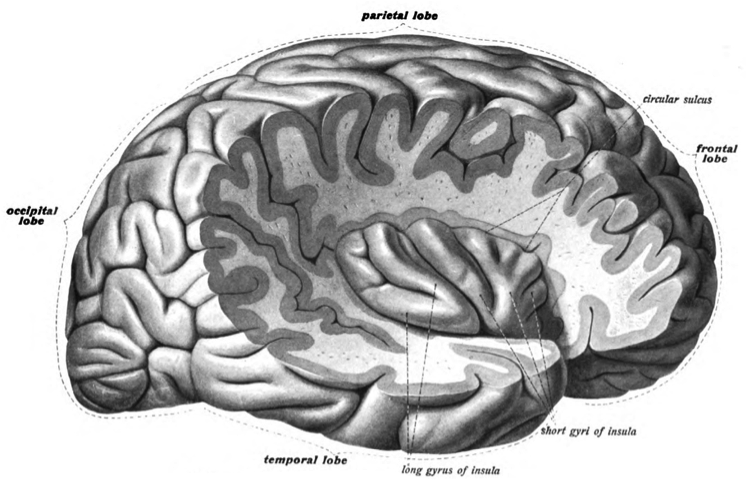
Though the cerebellum was located earlier, its function was omitted; it is responsible for motor coordination, posture and balance. The insula is deep region of the cerebrum near the junction of all four lobes, involved in the coordination of midline (head, neck, trunk) musculature. The insula, however, was only activated during overt singing or speech, not covert. This indicates that it is likely to be the area that mediates the control of the vocal tract musculature. The intrasylvian cortex, found inside the Sylvian fissure, the cleft that separates the temporal and frontal lobes, is an area of particular interest to those trying to find the neural pathway of singing. These intrasylvian structures are thought to be the centre of speech-motor coordination and the activation of the right-sided components during non-lyrical tune production indicates that they mediate the control of the vocal tract musculature during overt singing, in combination with the right cerebellum (Ackermann, Wildgruber & Riecker, 2006).
[/toggle]
[toggle title=”What is the next step in understanding how the brain turns metaphors into sound production?”]
 The lack of activation during covert singing highlights the intrasylvian cortex as a possible link between imagined singing and physical sound production. Its location along the borders of the temporal, parietal and frontal lobes places it in close proximity to the auditory cortex, Broca’s area, Wernicke’s area, and the motor and sensory cortices. However, just as more than one sense is activated when taking in a musical performance, the body requires synergy between various areas of the brain to coordinate any motor skill. The areas mentioned in the above sections are some of the many that combine to allow for the performance of vocal music.
The lack of activation during covert singing highlights the intrasylvian cortex as a possible link between imagined singing and physical sound production. Its location along the borders of the temporal, parietal and frontal lobes places it in close proximity to the auditory cortex, Broca’s area, Wernicke’s area, and the motor and sensory cortices. However, just as more than one sense is activated when taking in a musical performance, the body requires synergy between various areas of the brain to coordinate any motor skill. The areas mentioned in the above sections are some of the many that combine to allow for the performance of vocal music.
In the same way that combining a physiological and metaphorical approach provides teachers with multiple instructional tools, this information could possibly lead to the development of a more specialized teaching method. It is possible to apply this to other musicians, although the areas of the brain activated would have to be investigated separately, as different muscles and motor skills would be used when playing an instrument rather than singing. By studying the areas of the brain activated by various types of metaphors, as defined by Barten (Barten, 1998), it may be possible to pinpoint what types of metaphors would be most effective, and therefore useful, in the teaching process. In the same way that a number of elements combine to make a piece of music unforgettable, the brain combines several specialized areas to create pathways between metaphors used in teaching and the motor skills that become ingrained in our bodies over time.
[/toggle]
[toggle title=”References”]
Ackermann, H., Wildgruber, D., & Riecker, A. (2006). Singing in the (b)rain: cerebral correlates of vocal music performance in humans. In E. Altenmüller, J. Kesselring & M. Wiesendanger (Eds.), Music, motor control, and the brain (pp. 205-222). Oxford: Oxford University Press.
Barten. S.S. (1998). Speaking of music: The use of motor-affective metaphors in music instruction. Journal of Aesthetic Education, 32(2), 89-97. doi: 10.2307/3333561
Blank, S., Scott, S., Murphy, K., Warburton E., & Wise, R. (2002). Speech production: Wernicke, Broca and beyond. Brain 125, 1829-1838. http://dx.doi.org.proxy3.library.mcgill.ca/10.1093/brain/awf191
Clements. J. F. (2008). The use of imagery in teaching voice to the twenty-first century student (Ph.D dissertation). Retrieved from ProQuest Digital Dissertations. (UMI 3321469)
Denis, M. (2002). Can the human brain construct visual mental images from linguistic inputs?In A. M. Galaburda, S. M. Kosslyn & Y. Christen (Eds.), The languages of the brain (pp. 215-225). Cambridge, Mass: Harvard University Press.
Fields, V. A. (1972). How mind governs voice. The NATS Bulletin 22 (2), 2-10.
Freed, D.C. (2000). Imagery in early twentieth-century American vocal pedagogy. Journal of Singing, 56(4), 5-12.
Janata, P. (2001). Neurophysiological mechanisms underlying auditory image formation in music. In R. I. Godoy & H. Jorgensen (Eds.), Musical imagery (pp. 27-42). Lisse, Netherlands: Swets & Zeitlinger B.V
Kosslyn, S. M. (1994). Image and brain: the resolution of the imagery debate. Cambridge, MA: The MIT Press.
Kosslyn, S. M. (2005). Mental images and the brain. Cognitive Neuropsychology 22(3/4), 333-347. doi:10.1080/02643290442000130
Lane, S. J. (2002). Structure and function of the sensory systems. In A. C. Bundy, S. J. Lane & E. A. Murray (Eds.), Sensory integration: Theory and practice (pp. 35-70). Philadelphia: F. A. Davis Company.
Miller R. (1998). The reluctant student. Journal of Singing, 54(3), 41-43.
Miller, R. (2001). The syntax of voice technique. Journal of Singing, 58(1), 49-50.
Miller, R. (2006). The singing teacher in the age of voice science. In R. T. Sataloff (Ed.), Vocal health and pedagogy (pp. 199-202). San Diego, CA: Plural Publishing, Inc.
Purves, D., Augustine, G., Fitzpatrick, D., Hall, W., LaMantia, A-S., McNamara, J., & White, L.(Eds.). (2008). Speech and language. In Neuroscience (4th ed.)(pp. 687-706). Sunderland, Massachusetts: Sinauer Associated, Inc.
Purves, D., Augustine, G., Fitzpatrick, D., Hall, W., LaMantia, A-S., McNamara, J., & White, L.(Eds.). (2008) . Survey of human neuroanatomy. In Neuroscience (4th ed.)(pp. 815-842). Sunderland, MA: Sinauer Associated, Inc.
Riecker, A., Ackermann, H., Wildgruber, D., Dogil, G., & Grodd, W. (2000). Opposite hemispheric lateralization effects during speaking and singing at motor cortex, insula and cerebellum. NeuroReport 11 (9), 1997-2000.
Vennard, W. (1967). Singing: The mechanism and the technic. New York: Carl Fischer.
Zatorre, R. J. & Halpern, A. R. (2005). Mental concerts: Musical imagery and auditory cortex. Neuron 47(1), 9-12. doi: 10.1016/j.neuron.2005.06.013
[/toggle]
[toggle title=”Media”]
Drawing by Kirsten LeBlanc
All photos were sourced through creativecommons.org and are listed in order of appearance.
Thinking. geralt, Pixabay. Copyright 2015.
Cerebrum Lobes. Jkwchui, Wikimedia Commons. Copyright 2013.
Types of Cortical Areas. Open Stacks College, Wikimedia Commons. Copyright 2013.
Iris in the Right Eye of a Girl. Laitr Keiows, Wikimedia Commons. Copyright 2004.
Insular Cortex. Dr. Johannes Sobotta, Wikimedia Commons. Copyright 1908.
Puzzle. geralt, Pixabay. Copyright 2014.
[/toggle]
[/accordion]
Leave a Reply